Executive Summary
1. It is not surprising that Mata Menge yielded some hominin remains. Artefacts found there earlier for many years indicated hominin occupation.
2. Hominin fragments now described are undiagnostic taxonomically beyond saying that they are from a hominin. Small jaw/tooth size falls within the range of Pleistocene hominins.
3. They may be ancestors of any hominin living later on Flores, no specific link to just a putative species H. floresiensis can be made
4. These new finds do not change our interpretation of the hominin bone assemblage from Liang Bua site – a fragmentary sample of small bodied H.sapiens with one pathological individual.
This file records our research group’s preliminary responses to copies of two papers now in press at Nature:
- Brumm A, et al. (in press 2016) Age and context of the oldest known hominin fossils from Flores. Nature: doi:10.1038/nature17663
- van den Berg, G. (in press 2016) Homo floresiensis-like fossils from the early Middle Pleistocene of Flores. Nature: doi:10.1038/nature 17999
Background
The specimens reported in the above-cited papers were excavated in 2014, so the investigators have had access to them for approximately two years. We received embargoed copies of the papers four days ago, on June 3, 2016, so have had only a few days to review the material intended for publication.
We are pleased to learn of the additional artefacts and human gnathic remains from Mata Menge (MM). Beginning a decade ago (Jacob et al., 2006) our group has referenced this site repeatedly, noting the early date of material from it, and its importance in treating many matters concerning human evolution among Australomelanesian populations. The First Mariners by Robert Bednarik (the scholar who suggested to Mike Morwood that Flores presented promising sites for the study of human ancestry in this region) provides a broadly informative context for understanding the ancient webs of contact and dispersion among continental, island, and ocean populations going back through the time range that includes Homo erectus, s.l. populations in the broad sense, and their phyletically successive species, Homo sapiens.
In assessing the diagnostic meaning of the recently discovered Mata Menge specimens it is useful to begin with a key sentence in the abstract of the 2004 paper in Nature (Brown, et al., 2004) that first proposed the existence of the taxon referred to as “Homo floresiensis”:
“Here we report the discovery, from the late Pleistocene of Flores, Indonesia, of an adult hominin with stature and endocranial volume approximating 1 m and 380 cm3, respectively.”
Our group’s research, published in 2006 and subsequently, has shown that both of the above values are substantial underestimates. The best current value for the endocranial volume of LB1 (the only known skull attributed to “Homo floresiensis” from the Liang Bua site or any other) is 430 ml (cm3). More appropriate stature estimates for LB1 (the only individual for which femurs are known) are approximately 1.25 to 1.30 m, which overlap the statures of some living Homo sapiens on Flores and elsewhere. Our estimates of endocranial volume and stature have been confirmed independently.
The formal diagnosistic elements of the proposed species “Homo floresiensis” from Liang Bua (LB) are listed here (in left column) in order originally presented).
Right column notes assess whether the LB1 (and LB2) characteristics can be observed in the Mata Menge material now in press (van den Bergh, et al. 2016).
| Morphological characteristics listed in Brown, et al., 2004 (based on LB1 partial skeleton and LB2 isolated left P3 | Mata Menge gnathic remains Mandible SOA-MM4 Left I1, right P3, I1/2, left M1/2 Deciduous teeth (l and r dc) |
| small-bodied hominin | yes |
| bipedal | unobservable (but see “Note” below) |
| stature (body height) similar to, or smaller than, Australopithecus africanus | unobservable |
| endocranial volume similar to, or smaller than, Australopithecus africanus | unobservable |
| lacks masticatory adaptations present in Australopithecus and Paranthropus | probable |
| substantially reduced facial height and prognathism | unobservable |
| smaller postcanine teeth | probable |
| posteriorly oriented infraorbital region | unobservable |
| cranial base flexed | unobservable |
| prominent maxillary canine juga form prominent pillars, laterally separated from nasal aperture | unobservable |
| petrous pyramid smooth and with low relief | unobservable |
| styloid process absent | unobservable |
| without vaginal crest | unobservable |
| superior cranial vault bone thicker than Australopithecus and similar to H. sapiens | unobservable |
| supraorbital torus arches over each orbit and does not form a flat bar as in Javan H. erectus | unobservable |
| mandibular P3 with relatively large occlusal surface area, with prominent protoconid and broad talonid | unobservable |
| mandibular P3 with either bifurcated roots or a mesiodistally compressed Tomes’ root | unobservable |
| mandibular P4 also with Tomes root | yes, bifurcated root (but a polymorphic trait) |
| first and second molar teeth of similar size | unobservable |
| mandibular coronoid process higher than condyle | unobservable |
| ramus [of mandible] has a posterior orientation | unobservable |
| mandible without chin | unobservable |
| posterior inclination of [mandibular] symphysial axis | unobservable |
| posteriorly inclined alveolar planum with superior and inferior transverse tori | unobservable |
| ilium with marked lateral flare | unobservable |
| femur neck long relative to head diameter | unobservable |
| femur shaft circular and without pilaster | unobservable |
| high bicondylar angle | unobservable |
| long axis of tibia curved and the midshaft has an oval cross-section | unobservable |
Note: LB1 specimen attributed to “H. floresiensis” was reconstructed as quadrupedal by G. van den Bergh (2005) in Natuurwetenschap & Techniek. We are unaware of any subsequent changes published by Dr. van den Bergh of his earlier views but they are assuredly incorrect for LB1 (which undoubtedly was bipedal) and, by any reasonable scientific inference, to the fossils sampled from the Mata Menge hominin population.
Brief Comments:
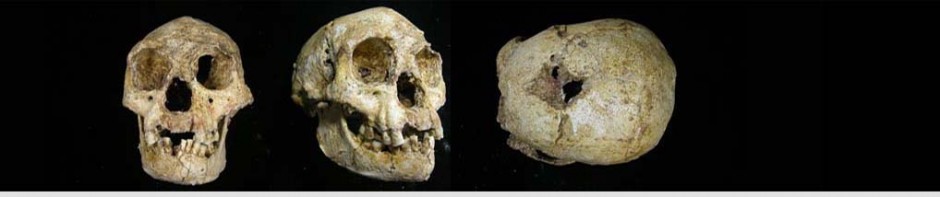
From the above preliminary comparisons it should be noted that the overlaps of the Mata Menge gnathic remains with those reported from Liang Bua are minimal. The most pertinent Liang Bua elements are the LB1 and LB6 mandibles, plus several isolated teeth. Sizes of Mata Menge teeth and the mandibular fragments are comparable (within ranges of individual variation) with various Pleistocene hominins. The molar crown SOA-MM1, that is small even in comparison to LB teeth, may be that of the second deciduous molar because its wear is slight. It shows proximal wear on the distal surface, but this does not argue against it being a deciduous tooth because permanent M1 lying just behind it erupts (now at age 6-7 years) while the second deciduous molar is still in place, and the second deciduous molar is lost only when replaced by a second premolar that erupts later (now at 10 years or a bit later).
The features shared in common between the two sites are principally overall body sizes inferred from the mandibles at the two sites. Reportedly, SOA-MM4 is smaller than LB1 and LB6. Size variation among humans in the Australomelanesian and surrounding regions should be considered in the context provided by the thesis of Dr. Bonita De Klerk (2012).
Curiously, the cranial fragment (SOA MM3) mentioned by Brumm et al. (2016) and listed in the Extended Data Table 1 by van den Berg et al. (2016) is neither pictured nor described in their paper. Since endocranial volume initially was a critical defining feature of “Homo floresiensis” and is disputed as being abnormally low due to developmental disruption, this omission is puzzling. The cranial vault bone of LB1 also has been stated incorrectly to be unusually thick since the initial description (Brown, et al. 2004), with this error repeated persistently since (e;g. Balzeau and Charlier, 2015). Consequently, even a small bone fragment might help correct this error as noted (Eckhardt, et al. 2014).
The new Mata Menge fossils do not help to resolve some of the most enduring questions that remain for bones from the LB site: endocranial volume, stature and limb proportions, taxonomic significance of chin presence or absence, and – perhaps still most important — whether the only relatively complete specimen, LB1 (which includes the only skull in the 62 bones it comprises from the site’s total of about 100 bones) is normal or abnormal. The most informative publications here include:
- Jacob, et al. (2006) PNAS
- Eckhardt, et al. (2014) PNAS
- Henneberg, et al. (2014) PNAS
- Eckhardt, et al. (2015) PNAS
The suggestion that comparisons among the Mata Menge and Liang Bua gnathic remains, in comparison to previously-known Homo erectus fossils, represent an “evolutionary reversal” overlooks an enormous body of work on size fluctuations in mammalian populations.
Similarly, there now is an existing body of knowledge about Homo erectus, s.l. populations. Many of these (Dmanisi) manifest small body sizes and extensive within-population variation; between-population comparisons of Dmanisi and other hominins with those from sites in other geographic regions (e.g. Nariokotome, Africa) show that, more than a million years ago, within and between-population differences approach some of those now found in our own species.
As they themselves have recognized, van den Bergh, et al. (in press 2016) provide data that are useful in countering the contention (by Argue and others) that the Liang Bua hominins probably are derived from unknown African early hominin ancestors). However, the case for such an African early hominin ancestor to the Liang Bua population never was plausible, and had been countered strongly by the extensive fossils discovered by Berger, Hawks, and their many colleagues in Dinaledi Chamber, South Africa (Eckhardt, et al., 2016).
Robert B. Eckhardt, Ph.D. Laboratory for the Comparative Study of Morphology, Mechanics, and Molecules Department of Kinesiology Huck Institute of Life Sciences Pennsylvania State University University Park, PA 16802 Phone: 814-237-1901
Maciej Henneberg, PhD, DSc, FAIBiol. Wood Jones Professor of Anthropological and Comparative Anatomy University of Adelaide, Medical School Adelaide, SA 5005, Australia +61-8-83135479, fax 83134398, mobile +61 414 885 983 Institute for Evolutionary Medicine University of Zurich Switzerland +41 44 635 01
